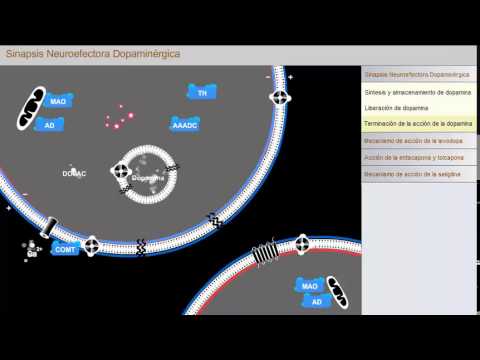
 La tirosina se transporta en la terminal neuronal de las neuronas dopaminérgicas y se convierte al producto intermedio L-dihidroxifenilalanina (DOPA, L-DOPA) por acción de la enzima tirosina hidroxilasa (TH). La DOPA se convierte a dopamina por acción de la descarboxilasa de L-aminoácidos aromáticos (AAADC, AAD). Más tarde, la dopamina se transporta a las vesículas de almacenamiento por el transportador 2 de monoaminas vesiculares (VMAT-2).
La tirosina se transporta en la terminal neuronal de las neuronas dopaminérgicas y se convierte al producto intermedio L-dihidroxifenilalanina (DOPA, L-DOPA) por acción de la enzima tirosina hidroxilasa (TH). La DOPA se convierte a dopamina por acción de la descarboxilasa de L-aminoácidos aromáticos (AAADC, AAD). Más tarde, la dopamina se transporta a las vesículas de almacenamiento por el transportador 2 de monoaminas vesiculares (VMAT-2).
La liberación de dopamina ocurre con la despolarización de la membrana celular presináptica, lo que causa la abertura de los conductos de calcio dependientes de voltaje, ocasionando la entrada rápida de Ca2+ en el citoplasma. El incremento de Ca2+ citoplásmico desencadena la fusión de las vesículas sinápticas con la membrana celular a través de la interacción de proteínas de membrana relacionadas con vesículas (VAMP) con proteínas asociadas con el sinaptosoma (SNAP). Las vesículas de fusión en la membrana celular ocasionan la exocitosis y liberación de dopamina a la hendidura sináptica. Una vez en la sinapsis, la dopamina puede interactuar con diversos receptores postsinápticos de dopamina, ocasionando una respuesta característica en la célula efectora postsináptica, lo que depende del tipo de receptor de dopamina presente.
Las acciones de la dopamina terminan por su recaptación en las terminales nerviosas presinápticas, donde se transportan a las vesículas de almacenamiento o se metabolizan. La dopamina también es captada y metabolizada por las terminales nerviosas postsinápticas. El metabolismo de la dopamina ocurre por acciones secuenciales de varias enzimas. En la terminal presináptica, la dopamina se convierte por acción de la monoaminooxidasa (MAO) y aldehído deshidrogenasa (AD) para formar 3,4-ácido dihidroxi fenilacético (DOPAC). Esta reacción también da origen a la formación de peróxido de hidrógeno (H2O2) que puede sufrir conversión espontánea a radicales libres de hidroxilo (OH) en presencia de hierro ferroso (Fe2+). Este radical reactivo de oxígeno puede contribuir a la degeneración de las neuronas dopaminérgicas, produciendo enfermedad de Parkinson en individuos susceptibles. La DOPAC formada se metaboliza a ácido 3-metoxi-4-hidroxi-fenilacético (también conocido como ácido homovaníllico, HVA) que es el principal metabolito de la dopamina en seres humanos. En neuronas postsinápticas, la dopamina se metaboliza por acción de la enzima catecol-O-metiltransferasa (COMT) a 3-metoxitiramina (3MT), que más tarde es metabolizada a HVA por acción de la monoaminooxidasa (MAO) y la aldehído deshidrogenasa (AD).
La levodopa (L-DOPA) se utiliza con fines terapéuticos para el tratamiento de la enfermedad de Parkinson porque se convierte principalmente a dopamina (DA) en las neuronas dopaminérgicas en el cuerpo estriado. La carbidopa por lo común se administra con levodopa porque antagoniza la descarboxilación prematura de la levodopa por la descarboxilasa de L-aminoácidos aromáticos (AAD) en la circulación periférica, con lo que se crean concentraciones encefálicas de levodopa y disminución de los efectos secundarios relacionados con la producción de dopamina en la periferia.
La entacapona y tolcapona inhiben el metabolismo de la levodopa administrada por vía oral en la circulación periférica por acción de la catecol-O-metiltransferasa (COMT). Cuando se administra simultáneamente con levodopa (L-DOPA), incrementa la cantidad de levodopa que alcanza el encéfalo. La tolcapona también actúa en el sistema nervioso central para inhibir la COMT y de esta forma reduce el metabolismo de levodopa y dopamina en el encéfalo.
En dosis bajas, la seligilina es un inhibidor irreversible selectivo de la isoenzima B de monoaminooxidasa (MAO-B), la forma predominante de MAO en el cuerpo estriado. La inhibición de MAO-B reduce el desdoblamiento de dopamina en las neuronas dopaminérgicas del cuerpo estriado, proporcionando tratamiento sintomático para la enfermedad de Parkinson.
3 jul 2015
![LIBRO | Fisiopatologia - Porth 7ed [ESPAÑOL] [PDF]](https://blogger.googleusercontent.com/img/b/R29vZ2xl/AVvXsEimjbL6LT_Z6d7Eu0E1yC6b2onCRoZtDwVpjEt35wdoXJs9ILNxAsP5_XNCdl5TapQ9TnvReb_aEPQgj20IfvVx5CuNL0aZathiFRcaxP_9WETriXKmL6CgcaWhnZVA9ef6k0qJpaWKynMJ/s72-w482-c-h591-no/1.jpg "LIBRO | Fisiopatologia - Porth 7ed [ESPAÑOL] [PDF]")
![LIBRO | Fisiopatologia - Porth 9ed [ESPAÑOL] [PDF]](https://blogger.googleusercontent.com/img/b/R29vZ2xl/AVvXsEi79OPTpVDjhWmNUjxiGBU-hC2JxPmpdfNR0wdyv_ftkeZO1PBSMcjwwpHQcUHSRnNdjMmiIsnKgh4yYE-0BlNrtrAax3dRTS3baRrXnt_PjmsEBzFWnkhvUOK2wA6SJYRR1ppNYHTBV6s/s72-c/porth.jpg "LIBRO | Fisiopatologia - Porth 9ed [ESPAÑOL] [PDF]")

![LIBRO | Rumack - Diagnostico por Ecografia 3ra Ed Tomo 1 [ESPAÑOL] [PDF]](https://blogger.googleusercontent.com/img/b/R29vZ2xl/AVvXsEj7TL7kFXNMJSZA8cAH-gH3RSpii2LbH2Au25HyqjWdfId5D_kF-_RZIMZeSdJJGkYRKwY1vBab9vS0xnatB1rvXHJ2kBvaBk4e868EHWZ_xj9SMcaH5Nvo1lr7glUHYuv3HMmtmpWNy0lS/s72-w306-c-h400-no/1.jpg "LIBRO | Rumack - Diagnostico por Ecografia 3ra Ed Tomo 1 [ESPAÑOL] [PDF]")
![LIBRO | Microbiologia Medica - Murray 7ma Ed [ESPAÑOL] [PDF]](https://blogger.googleusercontent.com/img/b/R29vZ2xl/AVvXsEisquvrRk6C-WfZIvx7QfTV-6WL0lqOuSBUJ589hpujlQi2rxQKdQzxDKZEJYqxzz4-Zug7lSQKw_BuhcuYK8KhIYD4V1k5PuDp1fe6T0mJdixY0snkxzHj8ahFMoTN3YyJC9j1yjee0_I/s72-c/Microbiologia.jpg "LIBRO | Microbiologia Medica - Murray 7ma Ed [ESPAÑOL] [PDF]")
![LIBRO | Goodman Gilman Las Bases Farmacológicas de la Terapéutica 12ed [ESPAÑOL] [PDF]](https://blogger.googleusercontent.com/img/b/R29vZ2xl/AVvXsEgUSn8akiMDV9L1ULfhk_ia031b25f9kF_BUy7_avmG16Y2QBGdHKC58Sms0MoNukK5SPnbFvS32LtxDfoxBDgaTC30vQTrXv7jvjUisAFjjXcH5Hm1Vv_AKrIZEBB9naUpfW1v-I2KosFz/s72-w455-c-h591-no/goodman.jpg "LIBRO | Goodman Gilman Las Bases Farmacológicas de la Terapéutica 12ed [ESPAÑOL] [PDF]")
![LIBRO | Dermatología - Fitzpatrick 7ed (Tomo 1) [ESPAÑOL] [PDF]](https://blogger.googleusercontent.com/img/b/R29vZ2xl/AVvXsEh_wYW6m9TkrkJ5XuFxXHMMm-_K1pEBgv_vqj3xatUwX4zLsFEnS3SzonOCyNZQrSzczFZQgiw34soMf6vd_wog1wBXqVOHSuWLyDQwxtq5UXZeMKfkCgpjwJdIjz76IHXVqBUSksFzW7Q/s72-c/Fitzpatrick.jpg "LIBRO | Dermatología - Fitzpatrick 7ed (Tomo 1) [ESPAÑOL] [PDF]")

![LIBRO | Neurología - Zarranz 5ed [ESPAÑOL] [PDF]](https://blogger.googleusercontent.com/img/b/R29vZ2xl/AVvXsEhkDSsoVvCseluT7N7anR_h1DK58lNVtVx5Vklbocu77zECw49tjocrQ-F8eKq6xjRbWE-JEaOTd5APklmRHSFK0s2QFT_I6r8P6MnXzCmcW0RSPkjrm1FyCtYGjeocDfFCDIi5OLdm1WXJ/s72-c/ZARRANZ.jpg "LIBRO | Neurología - Zarranz 5ed [ESPAÑOL] [PDF]")
